News Detail
 来源:2024-03-18 15:19:41
来源:2024-03-18 15:19:41
 浏览量:17462
浏览量:17462
银屑病(Psoriasis)俗称牛皮癣,是个体与环境相互影响诱发的免疫介导的慢性、复发性的、炎症性系统性的疾病,典型临床表现为局限或广泛的点疹、红斑、鳞屑和脓疱,没有传染性。银屑病发病范围较广,可影响世界各地区、各种族、各年龄段的人群,其中白人群中更为常见,且成人患病率高于儿童,[1]被世界卫生组织列为最严重的非传染性疾病之一。[2]银屑病大致可分为寻常型、脓疱型、红皮病型和关节病型4个大类,其中以寻常型最为常见,又可分为点滴状银屑病、斑块状银屑病(85%以上)、反向性银屑病。[3]虽然通常情况下银屑病是非致命性的,但银屑病可合并克罗恩病、代谢综合征和心血管系统疾病,增加患者的疾病负担,严重影响患者的生活质量,[4]并且银屑病对外貌的影响增加严重困扰患者的身心健康,并影响个人的个人、职业和婚姻生活。[5-6]
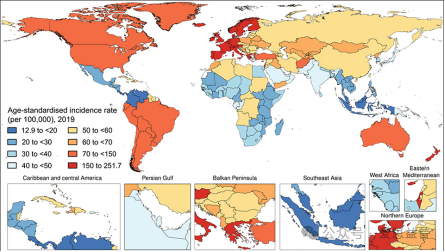
图一:银屑病年龄标化发病率[7]
银屑病发病机制较为复杂,可以由遗传因素[8]、免疫功能紊乱[9]和环境等因素相互作用共同导致。创伤、感染或药物治疗导致的表皮屏障破坏可能是银屑病发展的第一步。当表皮屏障遭到破坏后,机体的固有免疫被激活,引起一些细胞因子的释放,导致局部发生炎症。其中,以辅助性T细胞1(Th1)和Th17为主的免疫细胞群的在银屑病发展过程中发挥了重要的作用。当表皮屏障受损后,自身核苷酸释放到体液中与抗菌肽(cathelicidin antimicrobial peptide;CAMP)结合形成复合物,被以浆细胞样树突细胞(plasmacytoid dendritic cells;pDCs)为主的抗原递呈细胞(APCs)识别后,使得抗原特异性CD8+ T细胞激活、扩增并迁移到表皮,从而导致角化细胞释放可溶性因子,进一步增加局部炎症并刺激角化细胞增殖。pDCs释放的干扰素-α(IFN-α)和β(IFN-β)等炎症介质刺激髓样树突细胞(myeloid dendritic cells;mDCs)分泌额外的以肿瘤坏死因子(tumor necrosis factor;TNF)、白介素12(IL-12)、IL-23为主的促炎介质,这些促炎因子刺激以Th1、Th17和Th22为主的关键T细胞群活动,从而导致以表皮增生、棘层增生、血管增生、角化不全等特征性病变和一些额外的炎症细胞进入病变。[10]目前,免疫学和遗传学已经确定IL-17[11]和IL-23[12]是银屑病发病的关键驱动因素。

图二:银屑病发病机制[10]
01、TNF-α
肿瘤坏死因子-α(tumor necrosis factor α,TNFα)调节许多生物活性,如炎症、凋亡、趋化因子的产生和代谢。肿瘤坏死因子α三聚体(51kda)与受体结合,通过多种途径诱导寡聚化和信号转导,包括核因子κB1(NF-κB1)、丝裂原活化蛋白激酶(MAPK)信号级联、caspase-3和caspase-8凋亡[13-14]。通过激活NF-κB1途径,TNF-α诱导炎性细胞因子如IL-1β、IL-6、IL-8和粘附分子的快速产生[15]。因此,TNF-α与宿主对感染因子的防御有关,但也是促进和维持慢性炎症性疾病炎症的关键因素。目前已有多款TNF-α抑制剂获批临床。
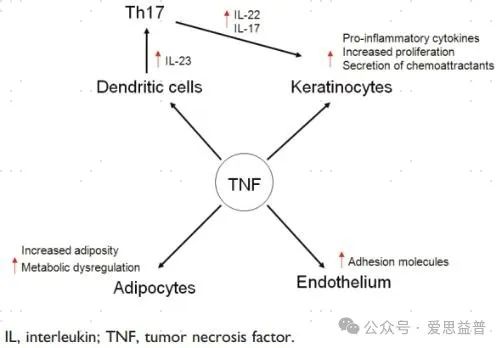
图三:TNF对各细胞的作用[16]
白细胞介素17(interleukin 17,IL-17)是Th17的主要效应因子,能够诱导上皮细胞、内皮细胞、成纤维细胞合成分泌IL-6、IL-8、G-CSF、PGE2,促进ICAM-1的表达的促炎因子。IL-17对内皮细胞的作用导致炎症和促凝血活性。当作用于上皮细胞和成纤维细胞时,IL-17会导致细胞因子和酶的产生。在单核细胞和树突状细胞上,IL-17通过增加促炎细胞因子的产生来促进炎症。[11]IL-17在银屑病中的主要靶点包括角质形成细胞、内皮细胞和先天免疫细胞,处于银屑病发病机制的中心位置。所以抑制IL-17会导致对银屑病发生和维持至关重要的信号通路的破坏。因此,靶向IL-17功能的生物制剂可快速显著改善银屑病和银屑病的皮肤和关节症状。

图四:IL-17对不同细胞靶点的影响[11]
03、IL-23白细胞介素23(interleukin 23,IL-23)与IL-17共同处于银屑病发展过程的中心位置。可以由多种多种细胞类型的细胞产生[17],它受Toll样受体信号转导的调控,并受TNF-α、IFN-γ和转录因子的增强[18-19]。IL-23受体(IL-23R)在记忆T细胞、自然杀伤细胞、中性粒细胞、肥大细胞、ILC和巨噬细胞上表达[19]。IL-23与其同源受体结合形成IL-23/IL-23R复合物,刺激ILC分化并触发CD4、CD8和γδT细胞合成IL-17和其他促炎细胞因子[20]。IL-23还诱导巨噬细胞产生TNF-α,在没有IL-17A的情况下刺激角质形成细胞增殖,并促进IL-23R的进一步表达,从而形成自扩增环[21]。
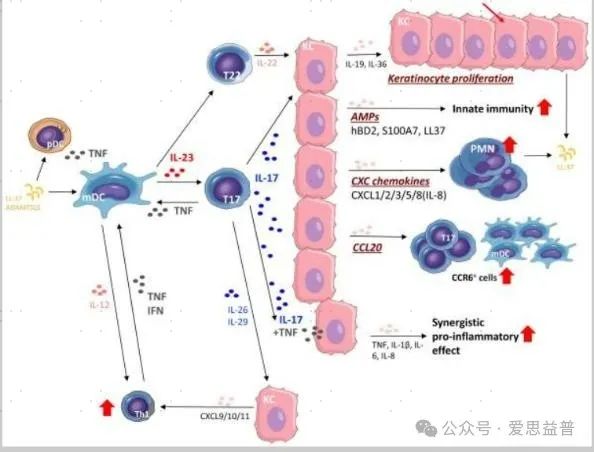
图五:IL-23/T17介导的对银屑病皮肤表皮角质形成细胞的影响[22]
04、JAKJanus集美信号转导和转录激活因子(JAK/STAT)是炎症和免疫反应的中枢调节因子,银屑病中重要的细胞因子IL-17、IL-23、TNF-α、IL-1、IL-22和IFN-γ均与JAK-STAT 通路相关[23],各种细胞因子与其各自的受体相互作用时,JAK的激活会导致STAT蛋白磷酸化和核易位,从而表达基因[24]。在银屑病中,参与树突状细胞的活化以及Th1和Th17细胞的分化的STAT1和STAT3的表达增加[25-26]。STAT3可通过IL-19、IL-36和 IL-22介导角质形成细胞增殖[27]。JAK抑制剂目前正在临床研究中用于口服和局部治疗银屑病[28-30]。目前,托法替尼、巴瑞替尼和芦可替尼这三种JAK抑制剂已在美国和欧洲获批用于银屑病临床。
05、STAT3STAT3是银屑病激活机制中的关键蛋白。首先,STAT3参与银屑病中关键细胞Th17细胞的分化。炎症反应中产生IL-6、IL-23等细胞因子,激活STAT3并形成IL-21/STAT3自分泌环路,引起STAT3的持续激活,上调RORγt的表达,促进Th17细胞的分化。其次,STAT3参与KC细胞的过度增殖与异常分化。Th17细胞产生的促炎因子IL-17、IL-22通过JAK2/AKT/STAT3信号通路激活KC细胞中STAT3的表达,进而促进KC细胞的过度增殖与棘层肥厚。第三,STAT3介导调节T(Treg)细胞低水平表达,造成机体免疫抑制功能下降,促进机体发生大量炎症反应激活银屑病的产生。最后,STAT3可以与血管内皮生长因子(VEGF)启动子结合,促进血管异常增生,此为银屑病另一重要病变特点。[26]
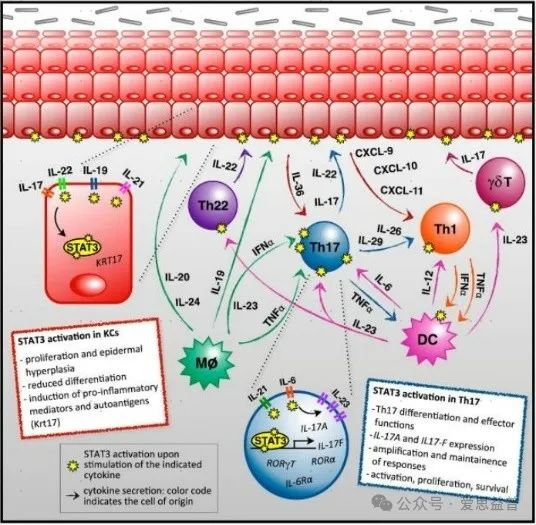
图六:STAT3在银屑病中的作用[26]
为寻找银屑病新靶点,目前已建立多种诱发动物银屑病模型的方式。
01动物选择小鼠作为实验动物有着成本低、饲养简单、成长周期短的优点,因此备受研究人员的青睐,目前基于小鼠的银屑病模型大致可以分为药物诱导模型、基因工程模型、细胞因子模型、异种移植模型四类。
02造模方法的选择2.1 自发型模型
自发产生银屑病样病变的小鼠模型是在自然条件下出现或经遗传育种筛选得到的由于基因突变产生的模型,该类较少,目前有缺皮脂突变鼠(asebia,ab/ab)、鱼鳞状皮肤突变鼠(ichthyosis,ic/ic),无毛突变鼠(hairless,hr/hr)和“鳞片状皮肤”突变鼠(flaky skin,fsn/fsn)等。由于目前发现的自发产生银屑病样病变的动物模型存在各种问题,尚未广泛使用。
2.2 诱发型模型
2.2.1 细胞因子注射模型
细胞因子注射模型是将细胞因子直接注射到小鼠皮肤或耳廓,从而造成局部皮肤出现银屑病样皮损。皮下注射的细胞因子包括IL-6、IL-21、IL-12和IL-23等。这些细胞因子可直接或间接激活Th17,产生IL-17从而导致局部出现银屑病样病变。这种方式诱导的模型易于操作、建模时间段且有着发病机制方面较为清晰,但由于是急性炎症一旦停止注射,症状即自行消退。
2.2.2异种移植模型
有研究者在严重免疫缺陷鼠中移植银屑病患者非病变皮肤后,移植皮肤出现银屑病部分病理变化,如棘层肥厚等。虽然该模型是从遗传、表型、免疫和致病过程最接近银屑病的动物模型,然而,因其需要获得大量银屑病患者组织,且需要快速收集和移植组织,再加上受体小鼠价格昂贵、饲养条件高,导致该模型并未得到广泛应用。
2.2.3基因编辑动物模型
采用基因编辑技术,过表达或敲除动物体内某种特定基因。K5.STAT3C转基因鼠模型、K5.TGFβ1转基因鼠模型、K14.AREG转基因鼠均是靶向角质形成细胞的基因编辑小鼠模型,使特定基因在角质形成细胞内过度表达,致细胞因子和趋化因子表达上调,从而使角质形成细胞发生增殖并促发炎性瀑布反应,最终产生银屑病样改变。。
2.2.4 药物诱导模型
药物诱导方式包括局部外用、真皮注射和口服,常用局部外用药包括咪喹莫特(Imiquimod, IMQ)、十二烷基硫酸钠和普萘洛尔等。其中以IMQ诱导的银屑病模型最为成熟、应用最广。
IMQ可以通过Toll样受体(TLR)7/8激活T淋巴细胞,从而产生白细胞介素、肿瘤坏死因子、干扰素等细胞因子,诱发皮肤出现皮肤增厚、角化不全等银屑病样的病变。该方法重复性高、造模方式简单,是研究银屑病病理机制及筛选药物最常用的模型之一。
爱思益普采用IMQ诱导小鼠产生银屑病样模型,可有效引起皮肤出现红疹、脱屑样病变,皮肤增厚和炎性浸润。数据展示如下:

参考文献
[1] Christopher E M Griffiths, April W Armstrong, Johann E Gudjonsson, et al. Psoriasis. Lancet[J]. 2021; 397(10281):1301-1315.
[2] Global report on PSORIASIS. World Health Organization. 2016.
[3] 中华医学会皮肤性病学分会银屑病专业委员会. 中国银屑病诊疗指南(2023版)[J]. 中华皮肤科杂志, 2023, 56(7):573-625.
[4] Komal Agarwal, Anupam Das, Sudip Das, et al. Impact of Psoriasis on Quality of Life[J]. Indian J Dermatol. 2022; 67(4):387-391.
[5] Beata Kowalewska, Barbara Jankowiak, Barbara Niedżwiecka, et al. Relationships between the acceptance of illness, quality of life and satisfaction with life in psoriasis[J]. Postepy Dermatol Alergol. 2020; 37(6):948-955.
[6] Ambra DI Altobrando, Giulio Vara, Federica Filippi, et al. The impact of psoriasis on marriage[J]. Ital J Dermatol Venerol. 2022; 157(3):235-239.
[7] Giovanni Damiani, Nicola Luigi Bragazzi, Chante Karimkhani Aksut, et al. The Global, Regional, and National Burden of Psoriasis: Results and Insights From the Global Burden of Disease 2019 Study[J]. Front Med (Lausanne). 2021; 16:8:743180.
[8] Nick Dand, Satveer K Mahil, Francesca Capon, Catherine H Smith, et al. Psoriasis and Genetics[J]. Acta Derm Venereol. 2020; 100(3):adv00030.
[9] Grace E Ryan 1, John E Harris 1, Jillian M Richmond. Resident Memory T Cells in Autoimmune Skin Diseases[J]. Front Immunol. 2021:12:652191.
[10] Jacqueline E Greb, Ari M Goldminz, James T Elder, et al. Psoriasis[J]. Nat Rev Dis Primers. 2016:2:16082.
[11] Andrew Blauvelt, Andrea Chiricozzi. The Immunologic Role of IL-17 in Psoriasis and Psoriatic Arthritis Pathogenesis[J]. Clin Rev Allergy Immunol. 2018;55(3):379-390.
[12] Lluís Puig. The role of IL 23 in the treatment of psoriasis[J]. Expert Rev Clin Immunol. 2017;13(6):525-534.
[13] J Simon C Arthur, Steven C Ley. Mitogen-activated protein kinases in innate immunity[J]. Nat Rev Immunol. 2013;13(9):679-92.
[14] Nina Van Opdenbosch, Mohamed Lamkanfi. Caspases in Cell Death, Inflammation, and Disease[J]. Immunity. 2019;50(6):1352-1364.
[15] Laura Barnabei, Emmanuel Laplantine, William Mbongo, et al. NF-κB: At the Borders of Autoimmunity and Inflammation[J]. Front Immunol. 2021:12:716469.
[16] John Yost, Johann E Gudjonsson. The role of TNF inhibitors in psoriasis therapy: new implications for associated comorbidities[J]. F1000 Med Rep. 2009:1:30.
[17] Andrea Chiricozzi, Paolo RomanellI, Elisabetta Volpe, et al. Scanning the Immunopathogenesis of Psoriasis[J]. Int J Mol Sci. 2018;19(1):179.
[18] Weicheng Liu, Xinshou Ouyang, Jianjun Yang, et al. AP-1 activated by toll-like receptors regulates expression of IL-23 p19[J]. J Biol Chem. 2009;284(36):24006-16
[19] Andrea Chiricozzi, Rosita Saraceno, Maria Sole Chimenti, et al. Role of IL-23 in the pathogenesis of psoriasis: a novel potential therapeutic target?[J]. Expert Opin Ther Targets. 2014;18(5):513-25.
[20] Tom C Chan, Jason E Hawkes, James G Krueger. Interleukin 23 in the skin: role in psoriasis pathogenesis and selective interleukin 23 blockade as treatment[J]. Ther Adv Chronic Dis. 2018;9(5):111-119.
[21] Andrea Chiricozzi, Paolo Romanelli, Elisabetta Volpe, et al. Scanning the Immunopathogenesis of Psoriasis[J]. Int J Mol Sci. 2018;19(1):179.
[22] Jason E Hawkes, Tom C Chan, James G Krueger. Psoriasis pathogenesis and the development of novel targeted immune therapies[J]. J Allergy Clin Immunol. 2017;140(3):645-653.
[23] Miguel Nogueira, Luis Puig, Tiago Torres. JAK Inhibitors for Treatment of Psoriasis: Focus on Selective TYK2 Inhibitors[J]. Drugs. 2020;80(4):341-352.
[24] Douglas A Harrison. The Jak/STAT pathway[J]. Cold Spring Harb Perspect Biol. 2012;4(3):a011205.
[25] A Hald, R M Andrés, M L Salskov-Iversen, et al. STAT1 expression and activation is increased in lesional psoriatic skin[J]. Br J Dermatol. 2013;168(2):302-10.
[26] Enzo Calautti, Lidia Avalle, Valeria Poli. Psoriasis: A STAT3-Centric View[J]. Int J Mol Sci. 2018;19(1):171.
[27] Shigetoshi Sano, Keith Syson Chan, Steve Carbajal, et al. Stat3 links activated keratinocytes and immunocytes required for development of psoriasis in a novel transgenic mouse model[J]. Nat Med. 2005;11(1):43-9.
[28] Lazaros I Sakka, Efterpi Zafiriou, Dimitrios P Bogdanos. Mini Review: New Treatments in Psoriatic Arthritis. Focus on the IL-23/17 Axis[J]. Front Pharmacol. 2019:10:872.
[29] Farzan Solimani, Katharina Meier, Kamran Ghoreschi. Emerging Topical and Systemic JAK Inhibitors in Dermatology[J]. Front Immunol. 2019:10:2847.
[30] Kata P Szilveszter, Tamás Németh, Attila Mócsai. Tyrosine Kinases in Autoimmune and Inflammatory Skin Diseases[J]. Front Immunol. 2019:10:1862.

业务咨询
北京
业务咨询专线:010-6780-9840
联系地址:北京市经济技术开发区科创十三街18号院锋创科技园16号楼
上海
业务咨询专线:010-6780-9840
联系地址:上海市浦东新区蔡伦路780号新技术推广大楼3E5O室
徐州
业务咨询专线:010-6780-9840
联系地址:江苏省徐州市云龙区淮海文博园 二号楼2层
贵州
业务咨询专线:010-6780-9840
联系地址:贵州省贵阳市南明区龙岭路50号 欧美医药产业园一期2号楼